





 and rabbit anti-beta tubulin Ab(AF7011) for 1 hour at 37°C. An AlexaFluor594 conjugated goat anti-mouse IgG(H+L) Ab(Red) and an AlexaFluor488 conjugated goat anti-rabbit IgG(H+L) Ab(Green) were used as the secondary antibody.
The nuclear counter stain is DAPI (blue).")
: control group, (right): Cytoflavin.")
 Real-time polymerase chain reaction (RT-PCR) showed that expression of lncRNA-UBOX5-AS1 was increased in ectopic endometrium tissues of endometriosis. (B,C) Representative western blotting analysis showing HIF-1α, E-cadherin, and vimentin protein expression in the different types of endometrium. β-actin was used as an internal loading control. *P<0.05; **P<0.01. lncRNA-UBOX5-AS1, long noncoding RNA UBOX antisense RNA 1; HIF-1α, hypoxia-inducible factor-1α.")
 Beclin-1 was measured by immunoblotting under normoxia and hypoxia. (b) Caspase-3 was measured by immunoblotting when incubated with HIF-1α inhibitor YC-1. The results indicate that hypoxia might induce cell chemoresistance through the HIF-1α signaling pathway. (c) Beclin-1 was measured by immunoblotting under normoxia and hypoxia. (d) Atg5 was measured by Western blotting and showed a similar trend to caspase-3. (e) HIF-1α was measured with Western blotting. The result revealed that HIF was involved in targeting apoptosis induced by autophagy.")
: Dual immunofluorescence staining for HIF-1α (red), CD68 (green) and DAPI (blue) indicated the co-expression of HIF-1α and CD68 (a macrophage marker) in human normal artery free of AS and atherosclerotic lesions from LE and CA with or without DM. (B-C): Compared with histologically normal artery, the protein levels of HIF-1α were upregulated in human atherosclerotic lesions from LE and CA with or without DM by western blot analysis. (D): Dual immunofluorescence staining for HIF-1α (red), CD68 (green) and DAPI (blue) in ApoE-/- mice fed with a HFD or normal diet demonstrated that the HIF-1α expression was present in macrophages, as indicated by HIF-1α immunoreactivity colocalized with CD68. (E-F): Compared with ApoE-/- control mice, the protein levels of HIF-1α were upregulated in HFD-fed ApoE-/- mice by western blot analysis. *P")
 Western blot analysis of the expressions of GLUT1, HK2, PKM2, LDHA and HIF-1α. Compared with NC group, the protein expression levels of GLUT1, HK2, PKM2, LDHA and HIF-1α in A549 cell line OV group were significantly increased. The protein expression levels of GLUT1, HK2, PKM2, LDHA and HIF-1α in H1299 cell line siRNA group were significantly lower than those in NC group. (B) After overexpression of AP000695.2, the uptake rate of 18F-FDG in OV group increased in A549 cells. After the expression of AP000695.2 was downregulated, the uptake rate of 18F-FDG in siRNA group in H1299 cells was decreased. (C) The content of lactic acid after overexpression of AP000695.2 was higher than that of the control group. Compared with the control group, the content of lactic acid was decreased after AP000695.2 was lowered. (D) The level of glycolysis, glycolytic capacity and glycolytic reverse were increased after overexpression of AP000695.2, while they were decreased after downregulation of AP000695.2 expression.")
 The effect of hypoxia treatment for 4 weeks on Cul5/TRAF6/NF-κB/HIF-1α/VEGF pathway expression in lung tissues of mice, and reverse effect of pevonedistat. (a) Western blot analysis of TRAF6/NF-κB/HIF-1α/VEGF pathway expression in N4W and H4W group and statistical analysis of the proteins relative expression normalized to GAPDH. (b) Western blot analysis of TRAF6/NF-κB/HIF-1α/VEGF pathway expression in N4W + DMSO, N4W + pevonedistat, H4W+ DMSO, and H4W + pevonedistat group, and statistical analysis of the proteins relative expression normalized to GAPDH. (B) The effect of hypoxia and/or Cul 5 treatment on TRAF6/NF-κB/HIF-1α/VEGF pathway expression in PAECs. Western blot analysis of TRAF6/NF-κB/HIF-1α/VEGF pathway expression in endothelial cells exposed to normoxia and hypoxia with or without Cul 5, and statistical analysis of the proteins relative expression normalized to GAPDH. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001. Student’s t test was used for statistical analysis in Figure 7A(a). Data are represented as the means ± SD, one-way ANOVA was used for statistical analysis in Figure 7A(b) and 7B.")
 Intracellular hypoxia in the kidneys of SI-AKI mice was detected by an immunofluorescence assay using a hypoxia probe (400x magnification). (c, d) Detection of HIF-1α protein expression in mouse renal homogenates by western blot. (e, f) The ROS level of kidney tissues was captured by CLSM at 24 h after LPS injection (dihydroethidium fluorescent staining, 400x magnification). (g, h) Intracellular hypoxia in the kidneys of SI-AKI mice after the knockdown of XO was captured by CLSM (400x magnification). (i, j) HIF-1α protein expression in SI-AKI mouse renal homogenates after downregulation of XO by western blot. (k, l) ROS levels in SI-AKI mouse renal tissue after the knockdown of XO were assessed using CLSM. Scale bar = 100 μm. ∗P < 0.05, ∗∗P < 0.01, and ∗∗∗P < 0.001 vs. control; #P < 0.05, ##P < 0.01, and ###P < 0.001 vs. LPS+Veh or LPS+Ctrl-shR (n = 10). Ctrl: control; Veh: vehicle; LPS: lipopolysaccharide; Feb: febuxostat; HIF: hypoxia-inducible factor; ROS: reactive oxygen species.")
 Representative immunohistochemical staining of Notch1 in AAV‐NC and AAV‐K3 mice post‐MI or Sham operation, including the remote area (RA) and border area (BA). (Scale bars = 20 μm). Notch1 expression levels were quantified and compared (n = 6). (B) qPCR analysis of Notch1 mRNA expression in AAV‐NC and AAV‐K3 mice post‐MI or Sham operation, including the remote area (RA) and border area (BA) (n = 4). (C) Representative Western blot images and quantitative analysis showing Notch1, Hes1, and Hif‐1α protein levels in CMECs. ns, not significant. Data were presented as mean ± SEM. **p")
 peptidase inhibitor, clade A, member 3C (Serpina3c) knockdown (3cKD) in adipocytes enhanced endoplasmic reticulum overoxidation and endoplasmic reticulum stress (ERS), and promoted the expression of pro-inflammatory cytokines and adipocyte apoptosis under lipotoxicity injury. (A) Western blotting of Serpina3c protein level in 3cKD 3T3-L1 adipocytes and its control LV3 group, as well as Serpina3c overexpression (3cOV) 3T3-L1 adipocytes and its control LV5 group. (B) Cell counting kit-8 (CCK8) assay measured the cell viability (%) of LV3 group and 3cKD group after 500 μM palmitic acid (PA)treated for 24 or 48 hours. (C-J) In these experiments, LV3, 3cKD, LV5, and 3cOV groups were treated by 500 μM PA for 48 hours. (C, D) The relative mRNA levels of indicated genes in 3T3-L1 adipocytes. (E) The protein levels of indicated genes in 3T3L1 adipocytes and quantification of the relative protein band density (n=3 for each group). (F) Hydrogen peroxide (H2O2) level in 3T3-L1 adipocytes was determined. (G) The protein levels of ERS makers in 3T3-L1 adipocytes and quantification of the relative protein band density (n=3 for each group). (H) Reactive oxygen species (ROS) level in 3T3-L1 adipocytes was detected. (I) The protein levels of mitogen-activated protein kinase signaling pathway in 3T3-L1 adipocytes and quantification of the relative protein band density (n=3 for each group). (J) Caspase-3 activity in 3T3-L1 adipocytes was determined. Data were presented as mean±standard error of the mean (n=5 for each group unless otherwise mentioned). NS, no significance; IL-6, interleukin-6; CCL2 or CCL5, C-C motif chemokine ligand 2 or 5; CXLC1 or CXCL10, C-X-C motif chemokine ligand 1 or 10; HIF1α, hypoxia-inducible factor 1α; Ero1α, endoplasmic reticulum oxidoreductase 1α; PDIA3 or PDIA4, protein disulfide isomerase family A member 3 or 4; GRP78, glucose regulated protein 78; CHOP, C/EBP homologous protein; p-eIF2α, phosphorylated eukaryotic initiation factor 2α; c-ATF6, cleaved activating transcription factor 6; XBP1S, spliced X-box binding protein 1; p-JNK, phosphorylated c-Jun N-terminal kinase; p-ERK, phosphorylated extracellular signal-regulated kinase.")
, eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1) and 70 kDa ribosomal protein S6 kinase (p70S6k), phosphorylated (p)-mTOR, p-4E-BP1 and p-p70S6k and hypoxia-inducible facto-1α; C: Quantification of protein expression using ImageJ. Data are presented as the mean ± SD (n = 3). aP < 0.05. bP < 0.01. cP < 0.001. HIF-1α: Hypoxia-inducible factor-1α; ns: No significance; RAPA: Rapamycin; ATP: Adenosine triphosphate; mTOR: Mammalian target of rapamycin; p-mTOR: Phosphorylated-mammalian target of rapamycin; 4E-BP1: Eukaryotic translation initiation factor 4E-binding protein 1; p-4E-BP1: Phosphorylated-eukaryotic translation initiation factor 4E-binding protein 1; p70S6k: 70 kDa ribosomal protein S6 kinase; p-p70S6k: Phosphorylated-70 kDa ribosomal protein S6 kinase.")
产品描述
*The optimal dilutions should be determined by the end user. For optimal experimental results, antibody reuse is not recommended.
*Tips:
WB: 适用于变性蛋白样本的免疫印迹检测. IHC: 适用于组织样本的石蜡(IHC-p)或冰冻(IHC-f)切片样本的免疫组化/荧光检测. IF/ICC: 适用于细胞样本的荧光检测. ELISA(peptide): 适用于抗原肽的ELISA检测.
引用格式: Affinity Biosciences Cat# BF8002, RRID:AB_2846221.
展开/折叠
ARNT interacting protein; ARNT-interacting protein; Basic helix loop helix PAS protein MOP1; Basic-helix-loop-helix-PAS protein MOP1; bHLHe78; Class E basic helix-loop-helix protein 78; HIF 1A; HIF 1alpha; HIF-1-alpha; HIF1 A; HIF1 Alpha; HIF1; HIF1-alpha; HIF1A; HIF1A_HUMAN; Hypoxia inducible factor 1 alpha; Hypoxia inducible factor 1 alpha isoform I.3; Hypoxia inducible factor 1 alpha subunit; Hypoxia inducible factor 1 alpha subunit basic helix loop helix transcription factor; Hypoxia inducible factor 1, alpha subunit (basic helix loop helix transcription factor); Hypoxia inducible factor1alpha; Hypoxia-inducible factor 1-alpha; Member of PAS protein 1; Member of PAS superfamily 1; Member of the PAS Superfamily 1; MOP 1; MOP1; PAS domain-containing protein 8; PASD 8; PASD8;
抗原和靶标
A synthesized peptide derived from human HIF1a.
Expressed in most tissues with highest levels in kidney and heart. Overexpressed in the majority of common human cancers and their metastases, due to the presence of intratumoral hypoxia and as a result of mutations in genes encoding oncoproteins and tumor suppressors. A higher level expression seen in pituitary tumors as compared to the pituitary gland.
- Q16665 HIF1A_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MEGAGGANDKKKISSERRKEKSRDAARSRRSKESEVFYELAHQLPLPHNVSSHLDKASVMRLTISYLRVRKLLDAGDLDIEDDMKAQMNCFYLKALDGFVMVLTDDGDMIYISDNVNKYMGLTQFELTGHSVFDFTHPCDHEEMREMLTHRNGLVKKGKEQNTQRSFFLRMKCTLTSRGRTMNIKSATWKVLHCTGHIHVYDTNSNQPQCGYKKPPMTCLVLICEPIPHPSNIEIPLDSKTFLSRHSLDMKFSYCDERITELMGYEPEELLGRSIYEYYHALDSDHLTKTHHDMFTKGQVTTGQYRMLAKRGGYVWVETQATVIYNTKNSQPQCIVCVNYVVSGIIQHDLIFSLQQTECVLKPVESSDMKMTQLFTKVESEDTSSLFDKLKKEPDALTLLAPAAGDTIISLDFGSNDTETDDQQLEEVPLYNDVMLPSPNEKLQNINLAMSPLPTAETPKPLRSSADPALNQEVALKLEPNPESLELSFTMPQIQDQTPSPSDGSTRQSSPEPNSPSEYCFYVDSDMVNEFKLELVEKLFAEDTEAKNPFSTQDTDLDLEMLAPYIPMDDDFQLRSFDQLSPLESSSASPESASPQSTVTVFQQTQIQEPTANATTTTATTDELKTVTKDRMEDIKILIASPSPTHIHKETTSATSSPYRDTQSRTASPNRAGKGVIEQTEKSHPRSPNVLSVALSQRTTVPEEELNPKILALQNAQRKRKMEHDGSLFQAVGIGTLLQQPDDHAATTSLSWKRVKGCKSSEQNGMEQKTIILIPSDLACRLLGQSMDESGLPQLTSYDCEVNAPIQGSRNLLQGEELLRALDQVN
研究背景
Functions as a master transcriptional regulator of the adaptive response to hypoxia. Under hypoxic conditions, activates the transcription of over 40 genes, including erythropoietin, glucose transporters, glycolytic enzymes, vascular endothelial growth factor, HILPDA, and other genes whose protein products increase oxygen delivery or facilitate metabolic adaptation to hypoxia. Plays an essential role in embryonic vascularization, tumor angiogenesis and pathophysiology of ischemic disease. Heterodimerizes with ARNT; heterodimer binds to core DNA sequence 5'-TACGTG-3' within the hypoxia response element (HRE) of target gene promoters (By similarity). Activation requires recruitment of transcriptional coactivators such as CREBBP and EP300. Activity is enhanced by interaction with both, NCOA1 or NCOA2. Interaction with redox regulatory protein APEX seems to activate CTAD and potentiates activation by NCOA1 and CREBBP. Involved in the axonal distribution and transport of mitochondria in neurons during hypoxia.
S-nitrosylation of Cys-800 may be responsible for increased recruitment of p300 coactivator necessary for transcriptional activity of HIF-1 complex.
Requires phosphorylation for DNA-binding. Phosphorylation at Ser-247 by CSNK1D/CK1 represses kinase activity and impairs ARNT binding. Phosphorylation by GSK3-beta and PLK3 promote degradation by the proteasome.
Sumoylated; with SUMO1 under hypoxia. Sumoylation is enhanced through interaction with RWDD3. Both sumoylation and desumoylation seem to be involved in the regulation of its stability during hypoxia. Sumoylation can promote either its stabilization or its VHL-dependent degradation by promoting hydroxyproline-independent HIF1A-VHL complex binding, thus leading to HIF1A ubiquitination and proteasomal degradation. Desumoylation by SENP1 increases its stability amd transcriptional activity. There is a disaccord between various publications on the effect of sumoylation and desumoylation on its stability and transcriptional activity.
Acetylation of Lys-532 by ARD1 increases interaction with VHL and stimulates subsequent proteasomal degradation. Deacetylation of Lys-709 by SIRT2 increases its interaction with and hydroxylation by EGLN1 thereby inactivating HIF1A activity by inducing its proteasomal degradation.
Polyubiquitinated; in normoxia, following hydroxylation and interaction with VHL. Lys-532 appears to be the principal site of ubiquitination. Clioquinol, the Cu/Zn-chelator, inhibits ubiquitination through preventing hydroxylation at Asn-803. Ubiquitinated by a CUL2-based E3 ligase.
In normoxia, is hydroxylated on Pro-402 and Pro-564 in the oxygen-dependent degradation domain (ODD) by EGLN1/PHD2 and EGLN2/PHD1. EGLN3/PHD3 has also been shown to hydroxylate Pro-564. The hydroxylated prolines promote interaction with VHL, initiating rapid ubiquitination and subsequent proteasomal degradation. Deubiquitinated by USP20. Under hypoxia, proline hydroxylation is impaired and ubiquitination is attenuated, resulting in stabilization. In normoxia, is hydroxylated on Asn-803 by HIF1AN, thus abrogating interaction with CREBBP and EP300 and preventing transcriptional activation. This hydroxylation is inhibited by the Cu/Zn-chelator, Clioquinol. Repressed by iron ion, via Fe(2+) prolyl hydroxylase (PHD) enzymes-mediated hydroxylation and subsequent proteasomal degradation.
The iron and 2-oxoglutarate dependent 3-hydroxylation of asparagine is (S) stereospecific within HIF CTAD domains.
Cytoplasm. Nucleus. Nucleus speckle.
Note: Colocalizes with HIF3A in the nucleus and speckles (By similarity). Cytoplasmic in normoxia, nuclear translocation in response to hypoxia (PubMed:9822602).
Expressed in most tissues with highest levels in kidney and heart. Overexpressed in the majority of common human cancers and their metastases, due to the presence of intratumoral hypoxia and as a result of mutations in genes encoding oncoproteins and tumor suppressors. A higher level expression seen in pituitary tumors as compared to the pituitary gland.
Contains two independent C-terminal transactivation domains, NTAD and CTAD, which function synergistically. Their transcriptional activity is repressed by an intervening inhibitory domain (ID).
研究领域
· Cellular Processes > Transport and catabolism > Autophagy - animal. (View pathway)
· Environmental Information Processing > Signal transduction > HIF-1 signaling pathway. (View pathway)
· Human Diseases > Cancers: Overview > Pathways in cancer. (View pathway)
· Human Diseases > Cancers: Overview > Proteoglycans in cancer.
· Human Diseases > Cancers: Specific types > Renal cell carcinoma. (View pathway)
· Human Diseases > Cancers: Overview > Central carbon metabolism in cancer. (View pathway)
· Human Diseases > Cancers: Overview > Choline metabolism in cancer. (View pathway)
· Organismal Systems > Immune system > Th17 cell differentiation. (View pathway)
· Organismal Systems > Endocrine system > Thyroid hormone signaling pathway. (View pathway)
文献引用

Application: WB Species: Mouse Sample:

Application: IF/ICC Species: Mouse Sample:
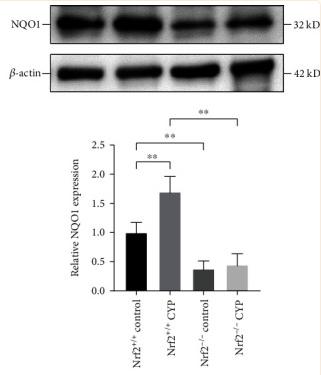
Application: WB Species: Mice Sample: bladder tissues
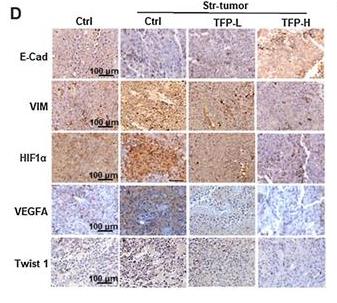
Application: IHC Species: mouse Sample: tumor cells
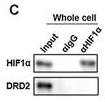
Application: WB Species: mouse Sample:
限制条款
产品的规格、报价、验证数据请以官网为准,官网链接:www.affbiotech.com | www.affbiotech.cn(简体中文)| www.affbiotech.jp(日本語)产品的数据信息为Affinity所有,未经授权不得收集Affinity官网数据或资料用于商业用途,对抄袭产品数据的行为我们将保留诉诸法律的权利。
产品相关数据会因产品批次、产品检测情况随时调整,如您已订购该产品,请以订购时随货说明书为准,否则请以官网内容为准,官网内容有改动时恕不另行通知。
Affinity保证所销售产品均经过严格质量检测。如您购买的商品在规定时间内出现问题需要售后时,请您在Affinity官方渠道提交售后申请。产品仅供科学研究使用。不用于诊断和治疗。
产品未经授权不得转售。
Affinity Biosciences将不会对在使用我们的产品时可能发生的专利侵权或其他侵权行为负责。Affinity Biosciences, Affinity Biosciences标志和所有其他商标所有权归Affinity Biosciences LTD.